生命の設計図からウイルスの正体まで、ミクロの世界を詳細に覗き見る魔法の顕微鏡「クライオ電子顕微鏡(cryo-EM)」。しかし、この強力なツールには、長年、研究者たちを悩ませてきた致命的な弱点がありました。それは、観察したい貴重なサンプルが、ほんのわずかな準備段階の操作でほとんど失われてしまうという問題です。このため、これまで多くの研究が断念されてきました。今回、ロックフェラー大学の日本人研究者らが、「磁石」を使ったシンプルなアイデアでこの課題を劇的に解決し、その応用範囲を大きく広げることに成功しました。

この新手法は「MagIC-cryo-EM」と名付けられ、磁気ビーズを用いて分子を所定の位置に保持することで、サンプルの完全性を保ち、サンプルロスを1000分の1にまで低減します。研究者たちは、2025年5月20日に学術誌『eLife』でその成果を発表しました。このオープンアクセスの論文のタイトルは、「MagIC-Cryo-EM, Structural Determination on Magnetic Beads for Scarce Macromolecules in Heterogeneous Samples(MagIC-Cryo-EM、不均一サンプル中の希少な高分子を対象とした磁気ビーズ上での構造決定)」です。
「MagIC-cryo-EMは従来法に比べてごく少数の粒子しか必要としないため、非常に希少なタンパク質や、作製・精製が困難なタンパク質など、より多様な分子の可視化に利用できます」と、筆頭著者であり、フナビキヒロノリ博士(Hironori Funabiki, PhD)が率いるロックフェラー大学染色体・細胞生物学研究室の元リサーチアソシエイトで、現在客員研究員を務めるアリムラヤスヒロ博士(Yasuhiro Arimura, PhD)は語ります。「ウイルスや他の感染症病原体の構造解析に特に有用かもしれません」。
革命的な洞察
2016年頃にクライオ電子顕微鏡用の革新的な電子カメラが「クライオEM解像度革命」をもたらしたとき、それは構造生物学に革命を起こしました。科学者たちはもはや、X線構造解析のために標的タンパク質を結晶化させる条件を見つけるのに何年も費やす必要がなくなりました。以来、クライオEMは、かつては解析不可能だった分子の内部構造を初めて可視化することを可能にし、生命の最も基本的なメカニズムのいくつかについて画期的な洞察を提供してきました。
クライオEMでは、標的粒子を液体バッファーに懸濁させ、微小な穴の開いたカーボンフィルム層を持つ小さな金のディスクに滴下します。このディスクをブロッティングして余分な液体を取り除き、液体エタンに投入して瞬間凍結させます。その結果、分子が詰まった薄い氷の膜で覆われたディスクが出来上がります――少なくとも、ブロッティング中にサンプルが失われなければ、の話です。その後、電子カメラが何百万もの低解像度の分子の2D画像を撮影し、ソフトウェアがそれらを組み立てて高解像度の3D構造モデルを構築します。
しかし、サンプルロスはクライオEMの唯一の欠点ではない、とアリムラ博士は言います。非常に小さなタンパク質の解析には本質的に不向きでもありました。「アルゴリズムは、S/N比が低いこれらの画像から小さなタンパク質を見つけるのに本当に苦労します」と彼は言います。「たとえ見つけられたとしても、それらを整列させて複合的な3D構造を形成するのは困難です」。
科学者たちがクライオEMの限界を克服したかったのは、まさしく彼ら自身の研究で繰り返しその壁にぶつかってきたからでした。フナビキ博士の研究室は、染色体遺伝の分子メカニズムに焦点を当てており、特に密に詰め込まれたヌクレオソームを研究対象としています。このヌクレオソームはその典型的な構造から、クライオEM技術開発の優れたベンチマークとして機能します。同時に、ヌクレオソームはゲノム機能の主要な制御標的の一つであるため、その構造がどのように認識され、変化するのかを理解することが重要です。ゲノムDNA配列の大きな多様性にもかかわらず、ほとんどのヌクレオソーム構造は、合成されたヌクレオソーム位置決め配列を用いて解析されてきました。アリムラ博士は、天然の条件下でヌクレオソームとその制御因子の構造を捉えたいと考えていました。
二つの問題の解決
サンプルロス問題の解決策は、驚くほど単純明快でした。アリムラ博士は、放射状に広がる「スペーサー」タンパク質のコロナで微小な磁気ナノビーズをコーティングし、標的分子を引き寄せて捕捉させました。そして、そのビーズを金のディスク上の穴の中に置き、底部に磁石を追加しました。このプロセスにより、標的分子は磁気ビーズに、磁気ビーズはグリッドに繋ぎ止められました。粒子がしっかりと固定されることで、はるかに少ないサンプルロスで安全にブロッティングが行えるようになったのです。
「従来のクライオEMでは、標的分子はバッファー中をランダムに漂っているだけでした」とフナビキ博士は言います。「しかし、私たちの方法では、分子がバッファー内に留まったまま余分な液体を取り除くことができ、すぐにガラス化できます。これにより、タンパク質の捕捉率が1000倍向上します」。
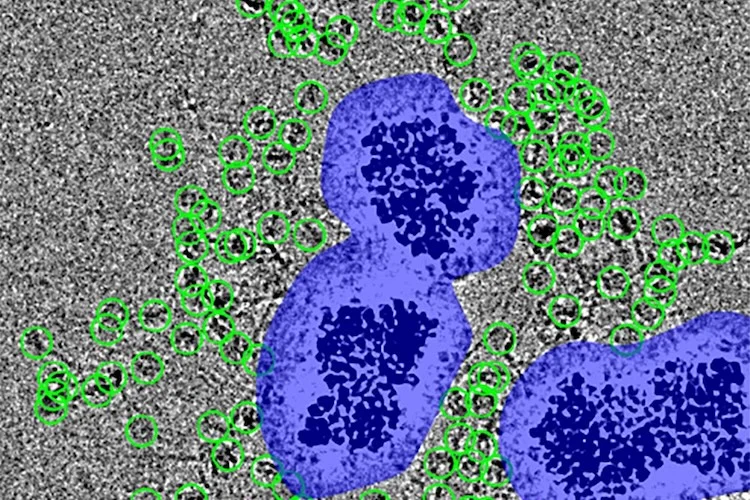
同時に、スペーサーはナノビーズによって導入される余分な視覚的ノイズも補償すると有村博士は付け加えます。「もし粒子が直接付着していたら、高出力の電子ビームが密度の高いビーズに当たって生じるホワイトノイズの中に埋もれてしまったでしょう」。
その結果、これらのタンパク質のハロー(光輪)を一つ一つ画像化し、3Dモデルを組み立てることが可能になりました。研究者たちはこの革新技術をMagIC-cryo-EM(Magnetic Isolation and Concentration:磁気による分離と濃縮)と名付けました。
アリムラ博士はまた、クライオEMの第二の限界、すなわち非常に小さなタンパク質を扱えない問題に対処する手法も開発しました。「DuSTER法(Duplicated Selection To Exclude Rubbish:ゴミを除外するための重複選択)」と名付けられたこのキュレーション手法は、S/N比が低い分子の画像を除外し、アルゴリズムが高品質な画像のみから3Dモデルを形成できるようにします。
今回の研究で、研究室はDuSTER法を用いて「NPM2」と呼ばれる分子の「開いた」状態を初めて画像化しました。シャペロンタンパク質と呼ばれるNPM2とその標的であるリンカーヒストンH1.8は、ヌクレオソームの組み立てに不可欠であり、「H1タンパク質は細胞分裂中に染色体を適切に形作る上で非常に重要です」とフナビキ博士は付け加えます。
「NPM2がどのようにH1に結合するのか、誰も知りませんでした」とアリムラ博士は言います。「それらはどのように互いを認識するのでしょうか?科学文献には多くの憶測や矛盾したデータがありました」。
DuSTER法で最もコントラストの低い画像を一掃した後、187,000枚以上の画像が残り、これらが組み立てられてNPM2とH1.8の史上初の3Dモデルが完成しました。様々な形で画像化されたこの構造は、明確な開いた状態と閉じた状態のバリアントを明らかにしました。H1.8は、NPM2タンパク質が開いた状態のときに結合することが判明しました。驚くべきことに、H1.8は他のタンパク質がゲノムDNAにアクセスする際のゲートキーパー(門番)としても機能していました。
将来の応用
これらの技術的進歩は、ウイルスを含む多種多様なタンパク質複合体の構造解析に利用できる、と舟引博士は言います。その目的のため、この研究はNIHおよびロックフェラー大学のSNF世界感染症研究所から支援を受けました。
「これらの技術は、例えばウイルス成分の構造解析に非常に役立つでしょう」とフナビキ博士は言います。「ウイルス成分は大量に作製するのが非常に難しいため、苦労して作製したものの、そのほとんどがブロッティング中に失われてしまうというのは、重大な問題でした」。
現在、フレッド・ハッチンソンがんセンターの助教として自身の研究室を率いるアリムラ博士は、これらのアプローチをさらに改良し、その応用を拡大しています。
「すでに世界中の多くの研究者から連絡をもらっています」と彼は言います。「私たちの技術が感染症研究にどのように貢献できるか、非常に楽しみにしています」。
画像:粒子(緑で囲んだ部分)は、ナノビーズ(紫で囲んだ部分)を囲むノイズから距離を置いて配置されている。(提供:フナビキ研究室)