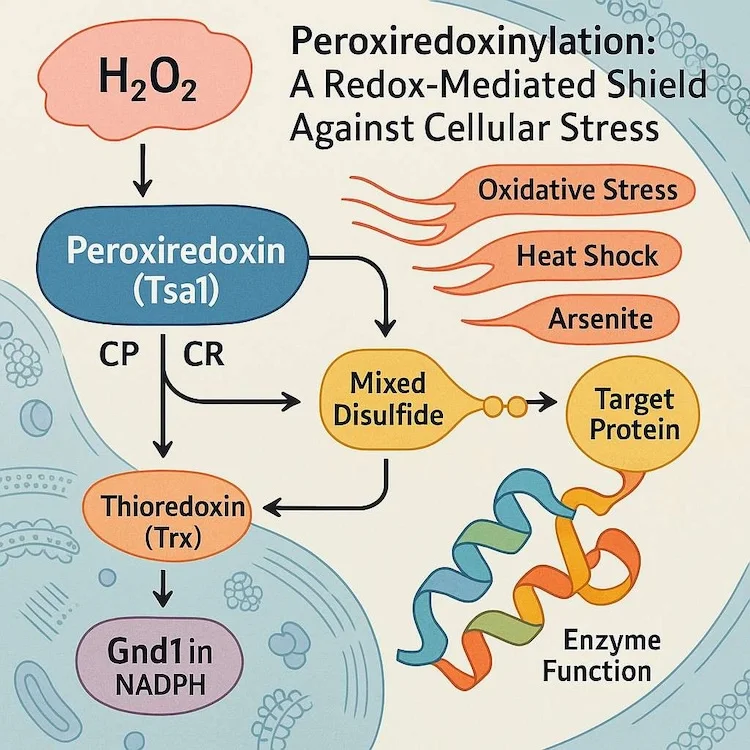
私たちの体を酸化ストレスから守る酵素「ペルオキシレドキシン」。この酵素の主な役割は、有害な活性酸素を除去することだと長年考えられてきました。しかし、最新の研究が、この酵素が持つ驚くべき「第二の顔」を明らかにしました。ストレスに晒された細胞内で、この酵素は重要なタンパク質に一時的に結合し、まるで盾のように保護する、これまで知られていなかった防御メカニズム「ペルオキシレドキシン化」の存在が突き止められたのです。この発見は、細胞がどのようにして過酷な環境を生き抜くのか、その理解を大きく塗り替えるものです。

2025年2月に「Cell Reports」誌に掲載された、サイゼンバッハー氏(Seisenbacher)らによる新しい研究「レドックスプロテオミクスはストレス保護におけるペルオキシレドキシン化の役割を明らかにする(Redox Proteomics Reveal a Role for Peroxiredoxinylation in Stress Protection)」は、真核細胞が環境ストレス下でタンパク質の機能を維持するのを助ける、これまで十分に評価されていなかったレドックスベースのメカニズムを提唱しました。この研究は、「ペルオキシレドキシン化」という新規の翻訳後修飾を特定しました。この修飾では、ペルオキシレドキシン酵素Tsa1が、細胞内の主要なタンパク質と共有結合性の混合ジスルフィド中間体(TIMDI: covalent mixed disulfide intermediates)を形成します。この修飾は、酸化的、熱的、化学的ストレッサーによって引き起こされる損傷から細胞を保護する上で、極めて重要な役割を果たします。
解毒作用を超えて広がるレドックス制御
ペルオキシレドキシン(PRX: Peroxiredoxins)は、過酸化水素を還元し、レドックス恒常性を維持するのを助ける普遍的な酵素です。その触媒機構やシャペロン機能は十分に文書化されてきましたが、今回の研究は、Tsa1が他のタンパク質に直接共有結合することを含む、レドックス感受性のタンパク質間相互作用ネットワークを明らかにしました。これらの混合ジスルフィド結合は、Tsa1のペルオキシダーゼ活性システイン(C48)に依存し、細胞のストレスに応答して形成されます。
研究チームは、Tsa1が広範な標的タンパク質と共有結合性のアダクトを形成することを発見しました。これらの結合はレドックス依存的で可逆的であり、Tsa1が活性酸素種の解毒剤としてだけでなく、レドックス制御されるタンパク質修飾因子としても機能することを示唆しています。このTsa1の新たな機能は、既知の過酸化駆動型シャペロン活性とは独立して作用し、Hsp70/Hsp104脱凝集システムやスルフィレドキシンSrx1介在の還元経路を必要としないようです。
ストレス下で増加する混合ジスルフィド中間体
野生型および変異型のTsa1を発現する酵母株を用いて、研究者らはTIMDIの形成が、中程度の過酸化水素暴露、亜ヒ酸処理、浸透圧ストレス、熱ショックなど、さまざまなストレス条件下で増強されることを示しました。対照的に、Tsa1の過酸化を誘導する高濃度の過酸化水素は、TIMDIの形成を妨げました。
興味深いことに、TIMDI形成は異常折り畳みタンパク質の蓄積の副産物ではありません。シクロヘキシミドやアゼチジン-2-カルボン酸など、翻訳を阻害したりタンパク質の異常折り畳みを促進したりする処理では、TIMDIは刺激されませんでした。これは、Tsa1が異常折り畳みされた新生鎖の救済機構として機能するのではなく、成熟した正常な折り畳み構造を持つタンパク質を標的にすることを示しています。
ペルオキシレドキシン化はストレスからの生存をサポートする
ペルオキシレドキシン化の生物学的重要性は、レドックスバランスを乱すことが知られている亜ヒ酸毒性に対する酵母の応答によって示されました。Tsa1を欠損する株や、TIMDI形成をなくす変異(例:C48S)を持つ株は、亜ヒ酸に対してより感受性が高くなりました。逆に、安定したTIMDI形成を促進するC171S変異体は、耐性を増加させました。TIMDIを抑制するために2番目の変異(Y78A)を導入すると、このストレス耐性は逆転しました。
これらの発見は、ペルオキシレドキシン化がストレス中に機能的に有益であり、プロテオームの完全性を脅かす条件下で細胞がタンパク質の活性と安定性を維持できるようにすることを示唆しています。さらに、この研究ではTIMDIとタンパク質のグルタチオン化パターンの重複が最小限であることがわかったため、ペルオキシレドキシン化は別のレドックス感受性修飾であるグルタチオン化とは独立して機能します。
プロテオームワイドなレドックス相互作用マップ
Tsa1のインタラクトームを包括的にマッピングするために、研究者らは還元条件下と非還元条件下でタンデムアフィニティー精製(TAP: tandem affinity purification)と質量分析を用いました。このアプローチにより、多数のレドックス感受性結合パートナーが明らかになり、C171S変異体は野生型Tsa1の211に対し、500以上の相互作用物質を捕捉しました。これらの結合の60%以上は非還元条件に特異的であり、共有結合性のシステイン介在結合を意味していました。
標的タンパク質は、タンパク質翻訳、リボソーム生合成、シャペロン活性、中心炭素代謝など、複数の生物学的プロセスに及びました。多くは5つ以上のシステインを持ち、ごく一部(1%)はシステインを全く持たず、これは全プロテオームと比較して有意に少ない割合でした。明確なアミノ酸モチーフやシステインの間隔パターンは特定されませんでしたが、システインリッチな領域の存在がTsa1との相互作用を促進するようです。
主要な標的の中に代謝酵素
Tsa1の最も興味深い標的の中には、グルコース代謝、トリカルボン酸(TCA)回路、アミノ酸生合成に関与する代謝酵素がありました。Cdc19(ピルビン酸キナーゼ)、Gpd1、Tdh3、Eno2、Ugp1、そして特にGnd1(6-ホスホグルコン酸デヒドロゲナーゼ)を含むこれらのいくつかは、生理的条件下で検出可能な安定したTIMDIを形成しました。共免疫沈降実験により、これらの相互作用が共有結合性でストレス誘導性であることが確認されました。
Gnd1-Tsa1の相互作用は特に重要であることが証明されました。Gnd1のシステイン460を変異させると、その主要な高分子量アダクトが消失し、酵素機能が破壊されました。野生型細胞では酸化ストレス時にGnd1活性が増加しましたが、この応答はGnd1C460S変異体やTsa1C48S株では見られず、ペルオキシレドキシン化がストレス誘導性の代謝適応に必要であることを示唆しています。
品質管理機構としてのペルオキシレドキシン化?
この研究はまた、ペルオキシレドキシン化が、異常折り畳みまたは構造的に不安定な酵素に対する品質管理の役割を果たすかどうかも探求しました。基質結合を損なうGnd1の変異(例:I366MおよびI366R)は、TIMDI形成の増加を引き起こしました。これは、Tsa1が触媒的に欠陥のある酵素にレドックス介在の品質管理タグとして結合する可能性を示唆しています。これらのアダクトはtsa1Δ株では見られず、Tsa1介在の修飾が特異的で制御されているという考えを裏付けています。
さらに、ペルオキシレドキシン化はタンパク質の発現レベルだけで決まるものではないようです。Tsa1と似たような存在量を持つTdh3のようなタンパク質でさえ、一貫してTIMDIを形成するわけではありませんでした。これは、システインのアクセスしやすさ、タンパク質の折り畳み状態、局所的なレドックス微小環境など、追加の制御層の存在を示しています。
チオレドキシンがTIMDIを解消する
ペルオキシレドキシン化の解消は、チオレドキシンシステムに依存することがわかりました。TRX1とTRX2の両方を欠失させると、ストレスがない状態でもTIMDIレベルが劇的に急増しました。これは、チオレドキシンが通常、これらの共有結合性アダクトの蓄積を防ぐように作用することを示しています。TRX1またはTRX2を回復させると、この表現型は救済されました。
インビトロでのアッセイにより、Trx1がジスルフィド交換を介して酸化されたTsa1部分を自身に転移させることで、Gnd1-Tsa1複合体を含むTIMDIを解消できることが確認されました。この解消経路は特異的であり、Hsp70、Hsp104、またはスルフィレドキシンの関与を必要としません。
広範な意味を持つレドックス翻訳後修飾
総合すると、これらの発見はペルオキシレドキシン化を、重要な生理学的結果を伴う、レドックス制御された可逆的な翻訳後修飾として位置づけます。これは、ストレス中に保護と調節のために成熟した機能的な酵素を標的にすることにより、標準的なレドックスリレーやタンパク質シャペロン経路とは異なります。この修飾は一過的で、酵素的に解消され、細胞のレドックス状態に合わせて調整されます。
著者らは、Tsa1機能の3層モデルを提唱しています:(1) 触媒サイクルによるH₂O₂の解毒、(2) 過酸化によって誘導されるシャペロン活性、そして(3) チオレドキシンによって可逆的にされる、標的タンパク質のストレス応答性ペルオキシレドキシン化。この柔軟なシステムにより、細胞は環境の変動に応じてタンパク質の活性、安定性、レドックス状態を迅速に調整できます。
考察
サイゼンバッハー氏らによる本研究は、ペルオキシレドキシン化を酵母における重要なストレス応答機構として位置づけ、ペルオキシレドキシンの機能的展望を広げるものです。レドックスプロテオミクス、変異導入、メタボロミクス、機能アッセイを組み合わせることで、著者らはTsa1が酸化的および環境的ストレスに応答して多数の標的タンパク質と可逆的な共有結合を形成するシステムを定義しました。これらの相互作用は主にペルオキシダーゼ活性システイン(C48)を介して起こり、一方、分解システイン(C171)の変異(C171S)は、これらの一時的な修飾を安定化させて捕捉するのに役立ちます。
重要なことに、Tsa1のレドックス感受性ネットワークは、以前に認識されていた過酸化水素の解毒やシャペロン活性における役割をはるかに超えて広がっています。ストレス中にTsa1プールの20%がTIMDIを形成するという発見と、500を超えるタンパク質相互作用物質が特定されたことは、ペルオキシレドキシン化がプロテオームレベルで広範な影響を持つことを示唆しています。この修飾は、代謝、翻訳、タンパク質恒常性の中心となる酵素に影響を及ぼし、細胞がストレス暴露中に必須機能を安定させるために使用する可能性のある一般的な戦略を示しています。
本研究はさらに、ペルオキシレドキシン化が新生タンパク質の合成や異常折り畳みに依存しないことを明らかにしました。代わりに成熟したタンパク質が標的とされ、この修飾は保護タグとして機能し、酵素活性を維持したり、有害なジスルフィド架橋を防いだりする可能性があります。このメカニズムは、特に過酸化やシャペロンの関与を引き起こさない中程度のストレス条件下で、古典的なレドックスリレーと並行して、あるいはその代わりとして機能する可能性があります。
TIMDIを解消する上でのチオレドキシン(Trx1およびTrx2)の役割は、本研究からのもう一つの大きな洞察です。これらの還元酵素がない場合、TIMDIは劇的に蓄積し、ペルオキシレドキシン化が動的で可逆的なレドックス制御メカニズムであるという考えを補強します。対照的に、Hsp70/Hsp104やスルフィレドキシンSrx1のような既知のシャペロンはTIMDIの解消に関与しておらず、この経路が以前に記述されたストレス応答とは異なることをさらに際立たせています。
特筆すべきは、Tsa1によって修飾される重要なシステイン残基としてGnd1C460が特定されたことで、ペルオキシレドキシン化が酵素活性の調節に直接関連付けられたことです。Gnd1C460S変異体がストレス誘導性の6-ホスホグルコン酸デヒドロゲナーゼ活性の増加を示せなかったことは、ペルオキシレドキシン化が主要な代謝経路を微調整できることを示しています。この機能は、多くが非常にストレス感受性の高い細胞プロセスに存在する他のTsa1相互作用物質にも及ぶ可能性があります。
結論
本研究は、ペルオキシレドキシン化を、細胞がレドックスの不均衡に適応することを可能にする、広範で生理学的に重要な翻訳後修飾として確立しました。Tsa1と広範な標的タンパク質との間の共有結合性でレドックス感受性の相互作用は、既知の解毒およびシャペロンシステムとは独立して機能する、プロテオーム保護の保存されたメカニズムを強調しています。
ストレス下で標的タンパク質を修飾し、チオレドキシン介在の解消を通じてそれらの回復を可能にすることで、Tsa1はレドックスセンサーとレギュレーターの両方として機能します。その相互作用は中心的な代謝酵素、特にGnd1に影響を与え、酸化的および環境的課題下での細胞の生存に直接貢献します。
これらの発見は、酵母におけるペルオキシレドキシンの役割を再定義するだけでなく、ヒトを含む高等生物にも同様のレドックス制御戦略が存在する可能性を提起します。哺乳類のPRDX1との共通の相互作用物質が特定されたことは、この考えを支持します。将来の研究では、ペルオキシレドキシン化によって支配される標的特異的な制御ネットワークが解明され、レドックスシグナリング、老化、疾患の病態における潜在的な意味が明らかになるかもしれません。
Tsa1がストレス下で標的タンパク質と可逆的な酸化還元結合を形成している図。